Introduction Zooplankton Biomass Zooplankton Species Composition Basin Scale Zooplankton Distributions Southwest coast of India and coast of Pakistan Somalia Region Central Arabian Sea Low Oxygen Zone and Zooplankton Distributions Problems of Sampling Zooplankton in Very Rough Seas
Monsoonal changes in wind strength and direction in the Indian Ocean result in dramatic seasonal changes in mixed-layer depth and phytoplankton concentration. During the SW monsoon, coastal upwelling occurs within 50 km of the Arabian peninsula. Further offshore, spatial gradients in the strength of the Findlater Jet during the SW monsoon cause Ekman upwelling and shoaling of the nutricline in the northwestern portion of the Arabian Sea. Coastal Zone Color Scanner images of chlorophyll-a just prior to and following the strong southwesterly winds of the summer monsoon show the development of intense blooms of phytoplankton from the coast of Arabia to 500 km offshore. The seasonal bloom of phytoplankton biomass is so extensive and of such high magnitude that it must be of ecological importance to the zooplankton species and populations of the northern Arabian Sea. We find nothing in the literature which describes or quantifies that ecology.
From the above account of the seasonal distribution of the biomass, it is evident that the areas adjacent to the Saudi* Arabian coast consistently show very high ranges of productivity during the prolonged SW monsoon period. A comparison of the biomass distribution with the surface thickness layer distribution makes clear that high biomass values overlap areas of no surface layer or regions of upwelling. In this respect the Saudi Arabian coast is unique....All these circumstances combine to make the Saudi Arabian coast the richest area for secondary production in the north Indian Ocean. [*Note: Saudi Arabia does not border the Arabian Sea; however Yemen and Oman on the Arabian Peninsula do].
As discussed in the section on physics, more recent studies (Bauer et. al., 1991; Smith and Bottero, 1977) suggest that high phytoplankton biomass off the Arabian peninsula result from two distinct SW monsoon upwelling regions: a coastal upwelling domain of ca. 50 km width, and a north central Arabian Sea upwelling domain (of ca. 500 km width).
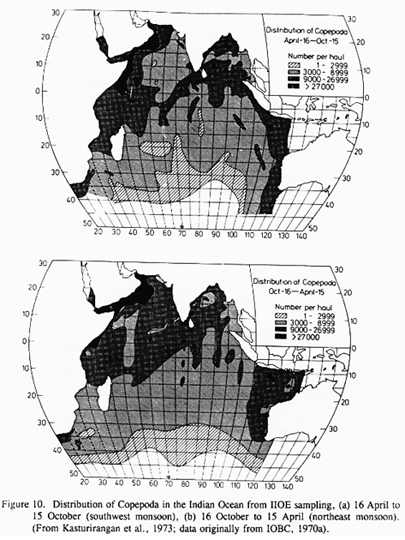
The zooplankton biomass of the Arabian Sea is largely comprised of copepods. The seasonal change in total copepod abundance is shown in Figure 10 (Kasturirangan et. al., 1973). Highest copepod densities occur along the northwestern border of the Arabian Sea. During April-October (SW monsoon) peak copepod abundances occur in the upwelling regions off Somalia and Arabia, but high densities also occur far offshore of the Arabian peninsula. During October-April (NE monsoon) copepods are abundant off the Arabian peninsula and in the Gulf of Oman, but are somewhat less abundant (compared to the SW monsoon) off Somalia. Unfortunately, the IIOE plankton collections have not been mined as extensively as one would hope at the species level. The distribution and abundance of only a few copepod species have been reported, notably, Gaussia princeps (Saraswathy, 1973a,b), Euchaeta spp. (Tanaka, 1973), Haloptilus spp. (Sarala Devi et. al., 1979), Pontella spp. (Pillai, 1975) and Candacia spp. (Lawson, 1977). Nor have the plankton samples from the IIOE been mined in another, potentially more valuable, way; namely, identification and enumeration of the entire copepod assemblage from samples collected within a defined region and season. Consequently, the reported results from the IIOE plankton investigations leave us ill-prepared to specify with any confidence which copepod species are dominant regionally or seasonally.
Of the large calanoid copepods the family Eucalanidae dominates the Arabian Sea with mean abundances of 3000 and 2250 per standard India Ocean haul (0-200 m) in the SW and NE monsoons, respectively (Stephen et al., 1992; these numbers are nearly identical to individuals per m2). The genera Euchaeta (1500/1900 for SW/NE monsoons) and Pleuromamma (270/1100 for SW/NE monsoons) are also abundant. Other represented taxa include the genera Lucicutia and Undinula, and the families Centropagidae and Temoridae. Of these the Centropagidae and Temoridae are coastal, Undinula is distributed in the region of the open-ocean upwelling, and Eucalanadae and Pleuromamma are found throughout much of the northern Arabian Sea (Stephen et al., 1992).
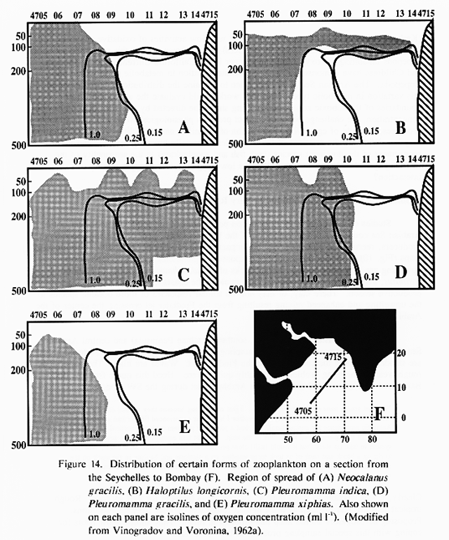
Species level analyses of the IIOE data are not complete and there is still some uncertainty in some of the systematics. For the copepods, Gopalakrishnan and Balachandran (1992) present the Scolecithricidae species from IIOE. Saraswathy (1986), Saraswathy and Krishnaiyer (1986), and Goswami et al. (1992) present data on Pleuromamma from various cruises. P. indica replace P. gracilis as the dominant species north of 10 deg N. This is consistent with P. indica making up between 21 and 95% of the myctophid diets in the surveys of Kinzer et al (1993). Saraswathy (1986) suggests that P. indica is tolerant to oxygen concentrations as low as 0.1 ml L-1 and is therefore adapted to the conditions in the northern Arabian Sea. Another species of interest is Calanoides carinatus which seems to dominate certain coastal upwelling systems at least episodically (Smith, 1982, 1984, 1992b). Calanoides is only found in 15 of the 340 IIOE stations sorted by Stephen et al. (1992). Only five of those stations are in the northern Arabian Sea; at one SW monsoon station 1175 individuals per standard haul were found. Two stations during the NE monsoon recorded 88 and 3 individuals.
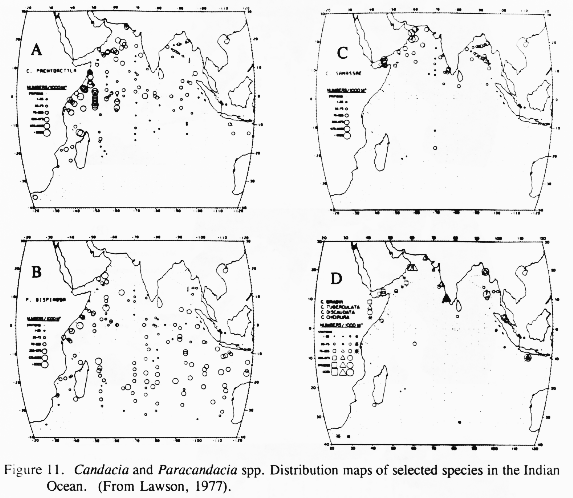
The most thorough, IIOE-based study of Arabian Sea copepods at the species level is Lawson's (1977) work on Candacia and Paracandacia. Lawson examined essentially the entire IIOE sample set covering the Indian Ocean to 35 deg S for this distinctive and well-studied genus. Distribution patterns for the Arabian Sea show both endemic species and species excluded from the region that are broadly distributed in the rest of the tropical Indian Ocean (Figure 11). Candacia samassae is endemic in all waters north of 10 deg N in both the Arabian Sea and Bay of Bengal. Paracandacia bispinosa is found essentially throughout the Indian Ocean, including the Bay of Bengal, but it is absent from the Arabian Sea north of 10 deg N (several exceptional occurrences are shown by Lawson right inshore off western Oman; Figure 11). These patterns suggest that the Arabian Sea is a distinctive pelagic habitat in some respect. Moreover, it has hydrographically distinct subregions to which mesoplankton populations respond by stock development or failure. Candacia bradyi is narrowly endemic in coastal upwelling along the Arabian Peninsula from the Gulf of Aden northeast to Oman. Although not reported off southwest India from the IIOE sampling, it shows up in SW monsoon samples off SW India (Madhupratap et al., 1992). It has no oceanic population at all. Candacia tuberculata is a coastal form whose range extends south along the Indian Peninsula from the Gulf of Oman to about Goa. It is not found elsewhere. Candacia pachydactyla, which is common across the equatorial belt becomes abundant in the Somalia Current and along the zone of the Findlater jet. However, it was present at only a few stations closer to the Arabian peninsula. On the other hand, Paracandacia truncata, which is common in tropical and subtropical waters worldwide, is common all over the Indian Ocean, including stock extensions nearly to the coast all around the Arabian Sea coast. Lawson's study of Candacia demonstrates that, as we find everywhere in every group of organisms, there are both habitat generalist and specialist species.
It can be expected that other copepod genera, when thoroughly studied will have both species endemic to the Arabian Sea and species with extended gaps in this region. For example, according to Rao et al. (1981), "The copepod Gaussia sewelli is endemic to the northern Indian Ocean particularly Arabian Sea and Bay of Bengal (Saraswathy, 1973). Another copepod showing limited distribution in the Arabian Sea and Bay of Bengal is Pleuromamma indica (Saraswathy, unpublished data)." (see also Saraswathy and Krishnaiyer, 1986).
Rao et al. (1981) report other species of zooplankton (beside the Candacia species studied by Lawson) that are either endemic to the Arabian Sea or have exceptionally large stocks there:
[A] preponderance of hydromedusae Aglaura hemistoma and Solmundella bitentaculata characterize the Arabian Sea water (Vannucci and Navas 1973). The cladoceran Evadne spinifera is found in the northeastern waters of the Arabian Sea (Della Croce and Venugopalan, 1973). Five out of the six species of the mysid belonging to the genera Paralophogaster are confined to the Red Sea and Arabian Sea while the species Siriella ionesi is limited to the Arabian Sea.
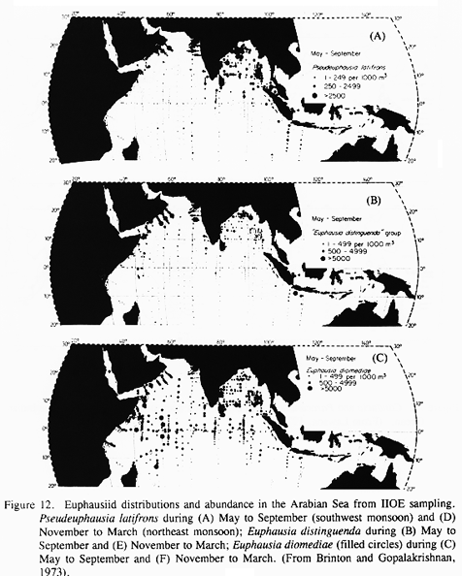
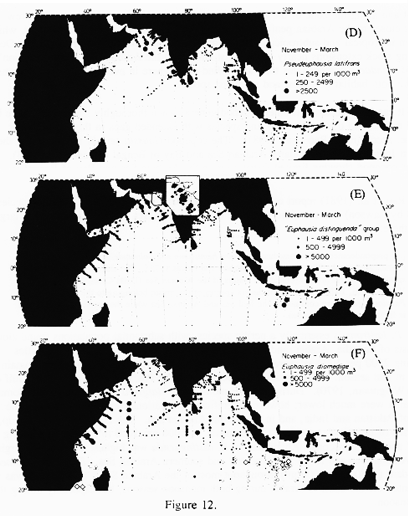
Pseudeuphausia latifrons, Euphausia distinguenda group, and Euphausia diomediae are the most abundant euphausiid species in the Arabian Sea (Brinton and Gopalakrishnan, 1973; Fig 12). P. latifrons is essentially a neritic species, with substantial populations found along the coasts of Somalia, Arabia, and India during the SW monsoon (Fig. 12). Densities exceeding 2500 per 1000 m3 were found in and offshore of the coastal upwelling domain off Arabia in that season. This species also occurs along the Pakistan coast and in the Gulf of Oman (Weigmann, 1970). During the NE monsoon, P. latifrons populations along Somalia and Arabia were much lower; however, significant populations were observed along the coasts of Pakistan and India, and in the central Arabian Sea. The E. distinguenda group, comprised of two similar species, was abundant (>5000 per 1000 m3) off Arabia during both the SW and NE monsoons. Abundance of E. distinguenda was highest around the margins of the Arabian Sea and lower in the central and southern Arabian Sea. Euphausia diomediae is abundant (>500 per 1000 m3) throughout most of the Arabian Sea. This species was well represented off the coast of Arabia during both monsoon seasons, but especially during the SW monsoon.
During the summer coastal upwelling, zooplankton species composition differed markedly between upwelled waters and offshore waters. In upwelling areas the dominant copepods were species of Eucalanus, Paracalanus, Clausocalanus, Temora, and Acrocalanus. Note the absence from these observations of capable workers of Calanoides carinatus, the "indicator species" for upwelling in the tropical Indian and Atlantic Oceans. In offshore, non-upwelled waters, Eucalanus attenuatus, Pleuromamma indica, and Pleuromamma abdominalis were dominant.
There have been few studies in the northern and eastern coastal regions which go beyond descriptions of zooplankton composition, abundance, and distribution. The dynamics of zooplankton populations in the region are virtually unknown (see Banse, 1984). The most extensive studies of plankton dynamics of the waters off SW India are those of Mathew and coworkers on the euphausiids. Prominent euphausiid species along the Indian coast are E. sibogae (a species in the E. distinguenda group), P. latifrons, and E. diomediae, in agreement with observations of the dominant species in the Arabian Sea basin as a whole. Maximum euphausiid abundances (>4000 per 1000 m3) along the southwest coast of India occurred during the August to October upwelling. This seasonal pattern was entirely caused by fluctuations of E. sibogae, which exceeded 4000 per 1000 m3 in August and October, but during other seasons remained less than 100 per 1000 m3. Conversely, populations of P. latifrons were greatest (200-500 per 1000 m3) during winter (December to February), and low throughout the remainder of the year. Breeding seasons of these three species were identified by examination of the seasonal occurrence of post-naupliar larval stages. Pseudeuphausia latifrons and E. diomediae have wintertime (December to April) breeding seasons while E. sibogae breeds during August to December (Mathew, 1983).
Copepods comprised >80% of the zooplankton enumerated from oceanic regions of the Arabian Sea during the NE monsoon (Madhupratap and Haridas, 1986). The northernmost samples (ca. 9 deg N) of that study were within the permanently oligotrophic region of the southern Arabian Sea. No single species accounted for greater than 10% of total copepod number. Copepod species diversity was fairly high (>40 species at some stations) even though small calanoid and cyclopoid copepods were undersampled because of the coarse (300 um) mesh of the net. Calanoid copepods were mostly typical subtropical (and small) copepods of the genera Paracalanus, Acrocalanus, and some Eucalanus. Calanoides carinatus, common in upwelling areas off South Africa (de Decker, 1973) and Somalia (Smith 1982, 1984), and observed off Oman (S. L. Smith, unpublished), was not observed in any oceanic samples during the NE monsoon season. Nor was C. carinatus mentioned by Sewell (1948) among the copepods of the John Murray Expedition of September 1933 to May 1934 in the Arabian Sea; however, this is not unexpected since they sampled off the Arabian peninsula during October to November, significantly after the SW monsoonal upwelling period.
Stephen (1984) reported on the distribution, abundance, and vertical distribution of copepods from the northern part of the Arabian sea (including Indian shelf and oceanic stations) in November. Dominant copepods in the surface layer at offshore stations (water depth >200 meters) were Eucalanus pileatus, E. subtenuis, Undinula vulgaris, and Cosmocalanus darwini. Below the mixed layer E. attenuatus, Pleuromamma indica, Haloptilus longicornis, and Lucicutia flavicornis were dominant.
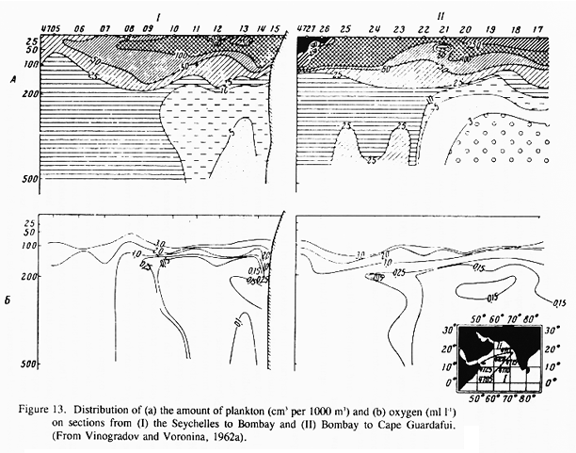
Vinogradov and Voronina (1962b) obtained vertically stratified plankton collections from the surface to 500 m along a transect from Bombay to Cape Guardafui during the NE monsoon season in 1959-60. Among the more common zooplankton along that transect were Euchaeta marina (50-100 m-2, 0-500m) and Undinula darwini (>100 m-2) among the copepods, and the large ostracod Pyrocypris sp. (>1000 m-2, 0-500m). The latter, a normally neritic species, was extremely abundant in the central Arabian Sea and relatively rare elsewhere in the western Indian Ocean.
Oxygen concentration in the eastern half of the northern Arabian Sea can be as low as 0.1 ml L-1. From studies in other low-oxygen environments (eastern tropical Pacific Ocean and elsewhere), most species avoid waters with oxygen contents below 0.1-0.2 ml L-1 (Longhurst, 1967; Judkins, 1980; Sameoto, 1986; Sameoto et al., 1987). There are, however, species which tend to reach maximum concentrations within oxygen minima at least part of the time. These include copepods in the eastern Pacific (Judkins, 1980; Sameoto, 1986) and a host of microzooplankton in anoxic fjords (Fenchel et al., 1990). Vinogradov and Voronina (1962a) show that low oxygen levels can influence zooplankton biomass distributions and alter species compositions in the Arabian Sea (Fig. 13 and Fig. 14). In other low-oxygen environments, fish have been observed to penetrate into waters that are anoxic to the point of hydrogen sulfide production (Baird et al., 1973). While the primary response to the low oxygen layer is undoubtedly behavioral modification, i.e. avoidance, it is clear that some organisms have means for coping with low oxygen environments.
Interfaces of oxygen minimum zones may be regions of high biological activity and aggregation sites for the whole trophic web from bacteria through mesopelagic fish (Wishner, 1991). There is evidence for dietary shifts to bacteria as the dominant food resource in these low oxygen habitats. They may also be regions where zooplankton organisms show biochemical or metabolic changes that affect their ability to live or take advantage of these regions. It is to be expected (Childress 1975, 1977) that plankton (and nekton, too) living continuously or for part of each day in suboxic waters will have strong capacity for anaerobic metabolism. This will be expressed (and can be studied) as high activities of glycolytic (Krebs cycle) enzymes and possibly correlated with low activities of oxidative enzymes. New work (Thuesen and Childress, in progress) indeed shows exactly this metabolic set up in copepods inhabiting the oxygen minimum layer in the California Current region. Theusen and Childress have demonstrated wide taxonomic variation in metabolic poise within the Copepoda. The Arabian Sea is an ideal place to examine the distributional correlations of this variation in metabolic type. Thus new work should evaluate the upper and lower boundaries of the suboxic zone, and sampling should be directed by real time data on oxygen concentration, the challenge being to connect previous physiological work with demographic processes, e.g. site of egg laying, distribution of juveniles, and mortality. A central question may be: What are the relations of the life cycles (especially breeding seasons) and the recruitment of the dominant species (both fish and zooplankton) to the seasonality of the various physical domains, and how does the low-oxygen waters at depth modify this interaction?
In both the coastal zone (Somalia, southern Arabian Peninsula) and central Arabian Sea there will be extreme problems for sampling and work at sea. Winds of 30 to 40 knots are standard for both the coastal jet and the Findlater jet. Waves are high. The scale of the rough weather sampling problem is probably quite severe. Heed this quote from A. deC. Baker who was on an IIOE cruise off the Arabian coast during the SW monsoon:
"On that cruise none of us were specialists. I just came along because they needed an extra pair of hands. It was the most consistently bad weather I have ever experienced [Baker had substantial Antarctic experience for comparison]. We had a particular problem with our vertical net hauls because of the up-and-down movement of the ship. The net was equipped with an accumulator system to take up slack, but it would [still] close prematurely when the wire went slack. We had to rig up a block and each of us took turns standing on a platform and hauling in the slack on the net. We were fishing five depths, and we each pulled on that line half an hour at a time. We were like bell-ringers." (Behrman, 1981, p. 72-73)
Clearly, it was possible to stand outside and ring bells by hand for half an hour. Rough tropical seas are less difficult to work in than rough seas in near freezing conditions. Proposing investigators should keep this problem in mind. They should show plans for coping with this special sampling problem in their proposals.
So, to achieve any significant zooplankton study in the Arabian Sea, we must be able to work fairly consistently in rough weather. Solving the rough weather sampling problem (for both fish and zooplankton) seems central to a successful U.S. GLOBEC study in the Arabian Sea. Essentially, it is necessary to equip a sizeable ship, that can move slowly in high winds and seas, to collect zooplankton samples with no personnel on deck. We need a crane or other mechanical arrangement for taking a ready sampler out of an interior, laboratory space, placing it overboard and lowering it to sample. It must then retrieve the sampler and return it to the laboratory for removal of the sample. For many kinds of work it will be necessary to be able to examine samples without too much wild rocking as the ship comes about in 40 foot waves. This may require one or more gimballed work stations including chair, working surface and microscope. Perhaps even a gyroscopically driven "gun table" would be justified; perhaps a small one could be obtained from the U.S. Navy. Each of these two requirements can be met by an engineering effort.
The Arabian Sea is stratified for several variables, with particular importance to zooplankton and fish attaching to the intense oxygen minimum layer from about 150 to 1500 m, particularly on the east side. Thus, characterization of the zooplankton community will require stratified sampling. Equipment for this that can be deployed and operated with no gear handlers on deck may be a difficult engineering problem. It is much simpler to deploy a single net for oblique or surface sampling from inside to outside, then over the side, down, and back. But that won't provide data on the vertical distribution of the fauna. For a multiple net sampler the most difficult part will be transfer of mesh fabric nets flopping wildly in a 50 kt wind back inside the ship. If we want to work effectively in this region, we must figure this out.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}