Justification of Target Zooplankton Species Calanoides carinatus Arabian Sea Euphausia Thalia democratica Ichthyoplankton and Fish Studies Approaches ConclusionThe implementation committee proposes that U.S. GLOBEC sponsor a study of modest scale that will take advantage of the ships and other logistics expected in the Arabian Sea region in 1994-1996. These include field programs of U.S. JGOFS, WOCE, ONR, and NOAA and imply the presence of three research vessels. The plan has been scaled so as to require very modest resources. Its returns will not necessarily be correspondingly modest, given the leveraging of resources that association with the other programs will provide.
While the problems of myctophid biology in the Arabian Sea are perhaps its most attractive feature for U.S. GLOBEC work (see below and Appendices A, B, and C), they are not particularly suitable for attack as a modestly priced enrichment of the U.S. JGOFS Arabian Sea Study. The logistic requirements for a detailed study of myctophid biology and distribution may be too different from those of U.S. JGOFS to be accommodated by a UNOLS vessel (however, for an alternative view see Appendix E). Of equal interest to U.S. GLOBEC is the extraordinary time and space variation of physical forcing (monsoonal reversals, coastal and open-ocean upwelling) and its potential effects on the pelagic ecosystem, including the zooplankton. A planktological study of great sophistication and detail can readily be mounted from a large UNOLS vessel. This can include net and specialized sampling to determine relationships among distribution and hydrographic features, field determinations of feeding rates, physiological and enzymatic experimentation, and more. A very rich body of information can be developed by a small number of U.S.-based investigators, particularly if cooperative programs develop with European, Omani, Pakistani, and Indian oceanographers.
The U.S. JGOFS Implementation Schedule (U.S. JGOFS Arabian Sea Process Study, Implementation Plan, May 1992, page 14) shows "cruises" aboard a large, otherwise U.S. JGOFS-specific, UNOLS vessel that are essentially intended as place holders for ONR/ARI and U.S. GLOBEC work for the following periods:
October 1994 July 1995 January 1995 October 1995The U.S. GLOBEC Steering Committee should request assignment of those cruises or additional ship-time for a U.S. GLOBEC study of the Arabian Sea. In order to maximize intercomparability of U.S. GLOBEC with U.S. JGOFS data, U.S. GLOBEC should carry out zooplankton and fish observations along the U.S. JGOFS station line (U.S. JGOFS Implementation Plan, page 5) which extends from the Omani coast at Cape Madraka (Ras al Madraka) to 8 deg N, 65 deg E (Figure 18). This would allow a U.S. GLOBEC observation set in the coastal upwelling regime off the Arabian Peninsula, through the upwelling hiatus landward of the Findlater jet, then through the oceanic upwelling regime, and finally out into persistently oligotrophic waters in the central Arabian Sea. Each expedition should be completed by occupation of a station in the NE Arabian Sea, in the location of U.S. JGOFS Station C (18 deg N 65 deg E; Fig. 18). This will allow detailed investigation of zooplankton and fish distribution and physiology in the region of most strongly developed, sharply defined suboxia at depth.
No substantive refitting of ships or basic scientific gear will be required for this work, over and above that required by U.S. JGOFS workers (CTD and rosette). However, individual proposing investigators will need to obtain sampling gear and move it to Oman for use in the U.S. GLOBEC cruises. Specific gear should be proposed by individual investigators. We recommend the following problems to the attention of the U.S. GLOBEC Steering Committee and to proposing investigators:
1) Detailed population analysis based on seasonal sampling for a suite of target zooplankton species. This should be accompanied by the most comprehensive measurements of functional ecodynamics for these species that the time available and timing of cruises permits. We recommend as target organisms:
2) A planktological reconnaissance of the Arabian Sea. This has been done, of course, by the John Murray Expedition of 1933, by the IIOE of the 1960's, and by the Soviets. However, by and large, a coherent, process-oriented view of the monsoonal Arabian Sea ecosystem has not emerged. Moreover, what is really lacking from prior investigations, including the Soviet literature, and what is particularly relevant to U.S. GLOBEC, is that the juveniles have not been studied, so that such basic issues as generation time, production, and seasonality related to the monsoon cycle cannot be addressed. In the opinion of the implementation committee, we haven't much idea which species of animals dominate the plankton in this region either as a whole or in specific sites at specific seasons. It is regrettable that we must recommend a study at this level after all the work in hand, but a basic study is needed.
3) Study of seasonality and general biology of midwater myctophids and ichthyoplankton of all types in the northern Arabian Sea. To what extent are larvae of Benthosema pterotum and other species variable in abundance and age structure with season? How fast do they grow and at what ages do they metamorphose into vertically migrating juveniles? At what depths do they live? What do they eat? Because of the traditional separation of ichthyological and general planktological expertise, actual study of fish larvae probably cannot be done by the same people working on the study of holoplanktonic target species described in (1). However, the same sampling techniques are required, and the samples can be cooperatively exploited by appropriate experts. Study of larval stages of the abundant Arabian Sea myctophids will set the stage for more extensive U.S. GLOBEC work (see FULL-SCALE Plan) on their biology when the opportunity arises. The fish working group at the Denver workshop recommended (see Appendix C) that a dedicated trawler vessel be requested for a comprehensive study of the distribution and ecology of midwater myctophids. Logistically, such a study cannot be conducted within the $1.5M constraint. Subsequently, the implementation committee has been convinced (see Appendix E) that much can be learned about the biology and distribution of adult myctophids using smaller, research-scale gear (1-10 m2 MOCNESS or equivalent) and acoustics from UNOLS vessels. We recommend that such studies be done, remembering that weather conditions may often prevent large-gear deployment.
Calanoides carinatus - This species was described by Kroyer in 1849. It is a 1.6 to 2.5 mm calanoid copepod distributed from the tropical Indo-Pacific as far east as New Zealand (Vervoort 1946, citing Farran 1929) westward to the coastal waters of Brazil and the Caribbean. In the Atlantic it is most abundant to the east, but it does occur all across the tropical zone (e.g., Bjornberg 1972). It is not reported from the eastern Pacific. As stated by Smith (1982) who worked in the Somalia Current during the SW monsoon upwelling, there is a "remarkable association of C. carinatus with upwelling...." This is true throughout its range, and Vives (1974) has suggested that in the vicinity of Cap Blanc (21 deg N, NW Africa) its presence is explicit evidence of upwelling (see also Weikert 1982). Extensive work in the Gulf of Guinea (Binet and Suisse de Sainte Claire 1975, Binet 1979, Mensah 1974a,b, Petit and Courties 1976, Petit 1978, 1982) and off South Africa (Borchers and Hutchings 1986, Peterson et al. 1990) shows the same thing all along the West African Coast. There is evidence that C. carinatus is associated with patches of upwelling that occur near islands and coastal features throughout the Malay Archipelago, off Taiwan, in the Tasman Sea, and elsewhere (Tranter 1977, Tranter et al. 1983). Occurrences of the species are associated with upwelling sites in the western Atlantic as well, particularly at Cabo Frio, Brazil (Valentin et al. 1987).
The seasonal phenology of C. carinatus is the same throughout its range. Intervals between upwelling seasons are spent in a fifth copepodite resting stage deep in the water column, well offshore. At the onset of upwelling, regardless of the local physical dynamics, the resting phase individuals are carried shoreward in the deep feeder flows to the upwelling. They arouse from their diapause and go through a number of rapid life cycles at intervals as short as 16 days, then return to depth and the diapause phase at the end of the upwelling season. No aspect of the environmental control or signalling involved in this alternation between active and resting life phases is known. Angel (1984) has suggested that perhaps "C. carinatus can only build up really large populations where there exists a quasi-stable eddy system" directly offshore of the upwelling zone. Smith (1982, pages 1350-1351) has calculated some of the parameters of population movement in just such a deep gyre in the NW Indian Ocean during the season spent submerged.
Some of the data of Binet are sufficient to support an estimate of secondary production attributable to C. carinatus off the Ivory Coast, but the calculations have never been made. It is certain, however, that huge production of biomass is attributable to this species over the African continental shelves, and it may be responsible for a major fraction of global copepod production. It is certainly an important component in nutrition of midwater fishes across the equatorial belt, although quantitative studies of is resting stock and transfer to fish remain to be made. In addition, there must be tropical pilchards and other coastal fish that feed on the growing stocks of C. carinatus, but those details also remain for study.
Several rearing studies have been done with C. carinatus, providing estimates of development time as a function of temperature and food availability. Hirche (1980) worked with the population in the upwelling region off northwest Africa (ca. 30 deg N); Tomasini and Petit (1977) succeeded with pre-feeding stages off Zaire; Borchers and Hutchings (1986) and Peterson and Painting (1990) reared individuals from the Benguela stock. Thus, it has been demonstrated that C. carinatus is readily amenable to experimental work. Moreover, the whole life history could be carried through several times in the space of a single oceanographic cruise.
Feeding rate studies have been done by Smith (1984) and Schnack (1982). Smith used the phytoplankton available in the ambient water; Schnack used cultures developed from local phytoplankton. Schnack's results were more extended and consistent. Smith's results showed a little less feeding, but were also reasonably consistent. Filtering rates were typically 325 ml female-1 day-1, a rate not unlike those for temperate calanids as determined by Gauld, Frost, Dagg, and a hundred others. Calanoides carinatus eats big diatoms and dinoflagellates readily in feeding experiments, and they were found by Schnack to have gut contents of small diatoms (Cyclotella sp., 15 X 5 um). Smith (1984, Table 4, p. 962) has produced a remarkable table for the Somali current, comparing C. carinatus stock size, and by implication feeding rates, with primary production. Where the copepod is abundant, right in the upwelling centers and upwelling-fed flow streams, it can eat half(!) of the primary production, which far exceeds any other system examined. Abundances in the Gulf of Guinea, where it constitutes >90% of mesozooplankton stock during the upwelling season, are higher than Smith's estimates for the Somali Current, so it may be even more important there.
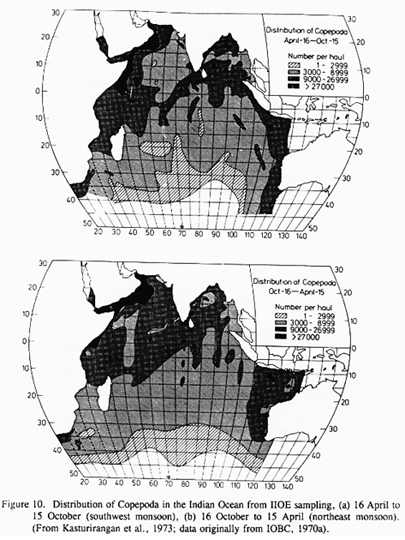
It is not fully established that C. carinatus is an abundant and recurring component of the copepod community during the upwelling season along the coast of the Arabian peninsula. However, we believe that it is because the previous sampling results (IIOE in particular) are so radically underreported (see however the large occurrences off Oman recorded from IIOE collections in a restricted area off Oman and, in lower abundance, off Pakistan (Stephen et al., 1992)). Also recent sampling (Sharon Smith, personal communication) has shown this species in substantial abundance at three stations off Oman during the SW monsoon. Continuity (Fig. 10) during the SW monsoon of the region of very high plankton biomass between the Somali coastal zone, where C. carinatus is known to be dominant, and the Arabian coast across the Gulf of Aden, suggests that the species is the upwelling season dominant at least as far east as the Gulf of Oman. The life cycle in the Arabian Sea must be tied somehow to the physics of the Arabian upwelling, since the species can be regarded as definitively absent from the long-lasting coastal upwelling off southwest India (cf. Madhupratap and Haridas, 1986; Madhupratap et al., 1992).
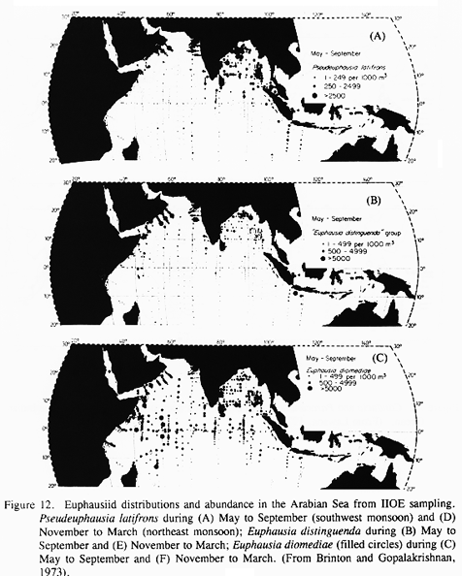
Arabian Sea Euphausia - The Arabian Sea has three species of euphausiid that appear to be particularly abundant there: Pseudeuphausia latifrons, Euphausia distinguenda, and Euphausia diomediae (Fig. 12). All three are also found in the Bay of Bengal. E. distinguenda (or a close relative) has a disjunct population in the eastern tropical Pacific, and E. diomediae is an Indo-Pacific tropical species with recurring high numbers in the Arabian Sea. We propose these species as a target group. Pseudeuphausia latifrons appears to be a very abundant species, but virtually all that we know of its biology comes from the distributional study of Brinton and Gopalakrishnan (1973). Repeating the information given above, it is essentially a neritic species, with substantial populations along the coasts of Somalia, Arabia and India during the SW monsoon. Although no samples are available, this species probably also occurs along the Pakistan coast and in the Gulf of Oman during the SW monsoon. During the NE monsoon, P. latifrons populations along Somalia and Arabia were much lower (although sampling off Arabia was sparse); however, significant populations were observed in the Persian Gulf, Gulf of Oman, along the coasts of Pakistan and India, and in the central Arabian Sea North of 15 deg N. No significant biological information whatever has been located about this species.
Distributions of the two Euphausia species (Brinton and Gopalakrishnan 1973) also extend to the shore, but they are more widely distributed across the Arabian Sea. They are on average more abundant than P. latifrons, and because they will be reliably available for study, they are excellent candidates for study of distributional shifts and life history adaptations in response to the seasonal variations of the monsoon. Their large body size and likely extended vertical migrations make them excellent candidates for study of metabolic adaptation of large zooplankton to suboxic conditions. Again, while we are certain that these species are consistently present and often abundant, we know essentially nothing of their detailed biology. Therefore, we propose that U.S. GLOBEC develop ecological and life-history information about these euphausiids.
Thalia democratica - Data on abundance of pelagic tunicates from the Arabian Sea are spotty in space and time, but several workers have reported very high densities of salps at several times of year (Sewell, 1953, Godeaux, 1972, Gjosaeter, 1984). Although several species are listed as common, Thalia democratica seems to be the most consistently abundant. Thousands of individuals were collected in the Gulf of Oman and northwestern Arabian Sea by the John Murray Expedition (Sewell, 1953), and Thalia occurred in 70% of the samples collected in the same region by Commandant Robert Giraud (Godeaux, 1972). The species is known to produce swarm concentrations associated with regions of upwelling world-wide (Berner, 1967, Heron, 1972, De Decker, 1973, Paffenhofer and Lee, 1987) and therefore should be well represented in the plankton of the productive inshore regions of the Arabian Sea. Thalia democratica are filter feeders capable of very high rates of ingestion and growth (Alldredge and Madin, 1982; Mullin, 1983, Heron and Benham, 1984) which enables them to respond rapidly to favorable environmental conditions (Paffenhofer and Lee, 1987). Preliminary biomass data from the Arabian Sea indicates that T. democratica may play a significant role in the seasonally varying zooplankton community of the mixed layer, but their role as grazers of primary production and competitors with crustacean consumers has not been evaluated. Because salps, and especially T. democratica, respond rapidly to changes in their environment, the implementation plan committee proposes that U.S. GLOBEC develop a better understanding of their population structure and dynamics in the seasonal upwelling regions of the Arabian Sea.
Zooplankton and fish sampling in the Arabian Sea during the height of especially the SW monsoon will require work in extreme sea states, certainly in sea state 6. This will require investigators to develop new, very rugged equipment, and it will probably preclude the use of classical, exposed nets altogether. For zooplankton, several solutions to this requirement can be imagined, including self-contained pumping-filtering systems in deployment packages resembling CTD-rosette systems. Development and deployment of such instrumentation is in line with U.S. GLOBEC's general goal of supporting improved technology for studies of pelagic ecology.
In addition to sampling with a newly developed, weather-hardened system, the zooplankton team should plan and propose a net-based sampling system for use in good weather and capable of filtering the large volumes required for capture and study of larval (and perhaps adult) fish. The system can be based on moderately coarse mesh (500 um) and can be built in a very rugged format for use in extreme sea conditions. The goal will be large samples of large plankton suitable for estimating stock numbers, size-frequency distributions, and biological characteristics of myctophid larvae. Since traditionally studies of ichthyoplankton and mesozooplankton are carried out by people with different interests and training, the zooplankton work should probably be divided among several investigators. However, cooperation in use of samples will be required, and it should be explicitly proposed by investigators.
The implementation committee leaves all details of approach and methodology to proposing investigators. The Arabian Sea provides an excellent ecological system for application of several of the new technologies for which U.S. GLOBEC has been providing support. Sonar methodology will have special applications for accurate estimation and spatial integration of the abundance of abundant myctophid fish. Specialized, high frequency sonar methods suitable for zooplankton studies may be ideal or certainly suitable for study of the target plankton species. All of the species populations are likely to be subject to subregional differentiation, and they will make good subjects for studies by modern, molecular methods in population genetics.
Some matters left to proposing investigators will eventually require negotiation and compromise among the successful candidates. These include station-spacing along the basic U.S. JGOFS line and duration and schedule of activities at each site. Possible plans include stations of several days duration at a series of locations. For example, on each cruise experimental and observation stations might be 1) over the shelf in the coastal upwelling, 2) in the gap between coastal and oceanic upwelling zones, 3) at the core of the Findlater jet (both when it is active and in other seasons), 4) offshore of the jet, 5) farther offshore in tropical oligotrophic waters, and 6) northeast of the main transect line in the most intensely suboxic region of the Arabian Sea. Intercalary sampling sites might be briefly visited during transit between these principal stations. Each main station could be occupied for 3-4 days on cruises of reasonable duration that fit within the overall U.S. JGOFS schedule for the ship. Consult Appendices A-D for additional information on potential approaches and logistical considerations.
{kind=link}
{kind=link}